진핵 생물의 재조합 DNA 기술에 대해 알아보자!
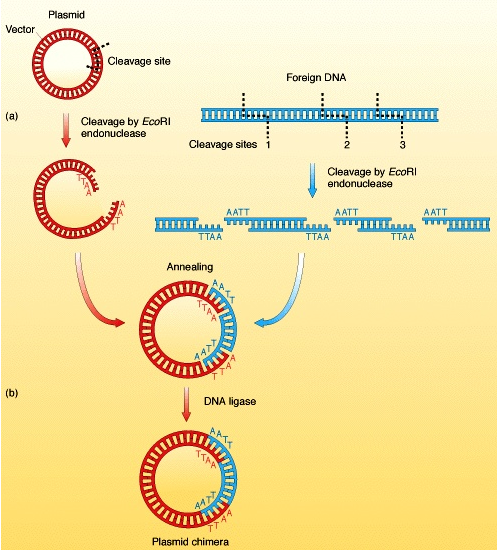
기술의 유전자 조작, 클로닝 및 발현은 박테리아제가 개발되었지만 현재의 진핵 생물 모델은 다양한 일상적인 형태로 받아드려 집니다. 진핵 생물의 게놈은 박테리아의 게놈 보다 더 크고 복잡하여서, 많은 양의 DNA 와 진 핵 생물의 다른 세포와 수명주기의 배열을 처리하기 위해 기술의 변형이 필요합니다. 예를 들어, 일부 진핵 생물 단백질은 박테리아에서 대량으로 쉽게 발현될 수 없으며, 진핵 생물 발현 시스템이 이용될 필요가 있습니다. 널리 사용되는 벡터 진핵 단백질 -expression 시스템에 도시된 바와 같이, 유전자가 삽입 배양 곤충 세포에서 높은 비율로 발현되는 바쿨로 바이러스 곤충 인도 진핵 생물 유전자는 박테리아 숙주에서 클로닝되고 서열화 되지만, 이러한 유전자를 원래 진핵 생물 숙주 또는 다른 진핵 생물, 다시 말해서 형질 전환 진핵 생물로 만들기 위해 다시 도입하는 것이 바람직합니다.
형질 전환된 진핵 생물
DNA는 변형, 주입, 바이러스 감염 또는 DNA 코팅 텅스텐 입자에 대한 충격과 같은 다양한 기술에 의해 진핵 세포에 도입됩니다. 원래 그 유기체에서 나온 DNA를 외인성으로 게놈에 삽입하면 상주 유전자를 대체 하거나 자궁외 삽입할 수 있습니다. DNA가 다른 종으로부터의 전이 유전자인 경우, 이소성 삽입, 진핵 세포에서 자율적으로 복제되는 벡터는 드물어서 대부분은 염색체 통합으로 따라오는 경로입니다.
식물 및 동물(인간포함)과 같은 진핵 생물의 유전자 도입 변형 가능성은 유전자형이 특정 실험에 적합하도록 유전자가 조작될 수 있기 때문에 연구에 대한 많은 새로운 접근법을 열어줍니다. 기본 연구의 예는 리포터 유전자를 사용하는 것입니다. 때로는 정상적으로 기능하는 조직에서 특정 유전자의 활동을 감지하기가 어렵습니다. 이 문제는 해당 유전자의 프로모터를 연결하여 우회할 수 있습니다. 리포터 유전자로 알려진 유전자의 코딩 영역으로 , 그 산물을 쉽게 검출할 수 있습니다. 문제의 유전자가 언제 어디서나 활성화될 때마다 리포터 유전자 적절한 조직에서 그 활동을 발표합니다. 또한, 식물, 동물과 곰팡이가 경제의 많은 부분을 형성하기 때문에 유전자 변형 "디자이너"유전자형이 응용 연구에서 광범위하게 사용되고 있습니다. 전이의 특히 흥미로운 적용은 인간 유전자 치료에 있습니다.
식물의 유전 공학
그들의 경제적 중요성 때문에, 식물은 개선된 품종의 개발을 목표로 하는 유전자 분석의 대상이 되어왔습니다. 재조합 DNA 기술은 이 기술에 의해 가능한 게놈 변형이 거의 무한하기 때문에 이러한 노력에 새로운 차원을 도입했습니다. 더는 번식은 주어진 종 내에서 변종을 선택하는 데 국한되지 않습니다. DNA는 이제 다른 종의 식물, 동물 또는 박테리아로부터 도입될 수 있습니다.
Ti 플라스미드
트랜스 제니 식물을 생산하기 위해 일상적으로 사용되는 유일한 벡터는 아기로박테리움투매파시엔스 (Agrobacterium tumefaciens)라 불리는 토양 박테리아로부터 유래된다. 이 박테리아는 크라운 담관 질환을 유발하며, 감염된 식물은 일반적으로 식물의 기저부 (크라운)에서 통제되지 않은 성장 (종양 또는 담즙 )을 생성합니다. 종양 생산의 핵심은 크게 (200-이다 KB ) 원형의 DNA를 플라스미드 - TI의 (tumor- 입회) 플라스미드. 박테리아가 식물 세포를 감염시킬 때, Ti 플라스미드의 일부 ( T-DNA라고 불리는 영역 )가 숙주 식물의 게놈으로 거의 무작위로 옮겨지고 삽입됩니다. 이 전달에 필요한 기능은 Ti 플라스미드의 T-DNA 외부에 있습니다. T-DNA 자체는 종양의 생성 및 오핀 이라고 하는 화합물의 합성을 포함하여 몇 가지 흥미로운 기능을 수행합니다. 오핀은 실제로 T-DNA의 지시에 따라 숙주 식물에 합성됩니다. 그런 다음 박테리아는 Ti 플라스미드의 오 핀 활용 유전자를 요구하여 자체 목적으로 오핀을 사용합니다. 두 가지 중요한 오핀은 노 팔림과 옥토 파인입니다. 2개의 별개의 Ti 플라스미드가 이들을 생산합니다.
Ti 플라스미드의 자연적 거동은 식물 벡터의 역할에 매우 적합합니다. 관심 있는 DNA가 T-DNA에 스플라이시될 수 있다면, 전체 패키지는 안정적인 상태로 식물 염색체에 삽입될 것입니다. 이 시스템은 실제로 이러한 방식으로 작동하지만 필요한 수정이 있었습니다.
Ti 플라스미드는 너무 커서 조작하기 어렵고 고유한 제한 부위가 거의 없어서 쉽게 작게 만들 수 없습니다. 결과적으로, 더 작은 중간 벡터는 초기에 관심 삽입물 및 재조합 , 복제 및 항생제 저항성에 필요한 다양한 다른 유전자 및 구분을 받습니다. 원하는 유전자 요소로 조작될 때 ,이 중간 벡터는 Ti 플라스미드에 삽입되어, 형질 전환으로 식물 세포로 도입될 수 있는 공적분 플라스미드를 형성할 수 있습니다. 중간 벡터를 수용할 Ti 플라스미드가 먼저 약의 독 화되고 즉, 종양 유전자 및 노 팔린 합성 유전자를 포함하여 T-DNA의 전체 오른쪽 영역이 결실되어 종양 형성이 불가능해져서 T-DNA 기능의 "불량" 측면이 됩니다. T-DNA의 왼쪽 경계를 유지하며 중간 벡터의 통합을 위한 교차 사이트로 사용됩니다. 중간 벡터는 다양한 독특한 제한 부위를 함유하는 편리한 클로닝 구분이 스플라이시 되었습니다. 관심 유전자는 그림 13-16 의의 사이트에 삽입되었습니다. 선택 가능한 박테리아 유전자 ( SPC R)도 벡터에 접합) 자격조건 티로 마이신 내성, 식물에서의 발현을 위해 조작된 박테리아 카나마이신-내성 유전자 ( Kan R ); 및 T-DNA의 두 세그먼트. 하나의 구분은 노파 린 합성 유전자 (NOA) + 우측 T-DNA 경계 서열을 보유합니다. 두 번째 T-DNA 구분은 왼쪽 경계 근처에서 제공되며, 해제된 Ti 플라스미드에 유지된 왼쪽 영역의 상동 부분과의 재조합 섹션을 제공합니다. 중간 벡터가 무장 해제된 Ti 플라스미드를 함유하는 아기로 박데 리움 세포 내로 도입된 후 플라스미드 재조합체 (공적 본체)는 자격조건 티로 마이신 상에 도금함으로써 선택될 수 있습니다. 중간체 벡터가 아기로 박데 리움에서 복제될 수 없어서 선택된 박데리아 군생은 Ti 플라스미드만을 함유할 것입니다.
클로닝된 DNA의 발현
DNA에 클로닝 T-DNA는 연구자가 대상 식물에 삽입하기를 원하는 모든 DNA가 될 수 있습니다. 특히 현저한 외래 DNA T-DNA를 사용하여 삽입된 인 유전자 위한 효소 반딧불이 루시퍼로부터 분리됩니다. 이 효소는 루시페린이라는 화학 물질과 ATP의 반응을 촉매합니다. 이 과정에서 빛이 방출되어 반딧불이 어둠 속에서 빛납니다. 루시패라제 유전자를 발현하는 형질 전환 담배 식물도 루시페린 용액으로 물을 뿌릴 때 어둠 속에서 빛날 것입니다. 이러한 조작은 조명이 필요 없는 크리스마스트리를 만들기 위한 기술을 개발하려는 시도처럼 보이지만 실제로 루시패라제 유전자는 개발 중에 유전자의 기능을 감시하는 리포터로 유용합니다. 즉, 업스트림 프로모터 임의의 관심 유전자의 서열을 루시퍼가제 유전자에 융합시키고 T-DNA에 의해 식물에 넣을 수 있습니다. 루시패라제 유전자는 일반적으로 조절되는 유전자와 같은 발달 패턴을 따르지만, 루시패라제 유전자는 조절 순서에 따라 다양한 시간 또는 다양한 조직에서 빛을 발함으로써 그 활성을 현저하게 발표할 것입니다.
'생명과학' 카테고리의 다른 글
| 말라리아(malaria) 감염 경로, 증상, 치료법에 대한 이해 (0) | 2020.07.18 |
|---|---|
| 구조 유전체학에 대해 이해하기!! (0) | 2020.07.18 |
| 바큘로 바이러스(baculovirus)의 발견!! 분류법 및 진화에 대한 이해 (2) | 2020.07.17 |
| 유전자 스위치란 무엇일까? (0) | 2020.07.17 |
| 염색체 DNA와 염색질 섬유에 팩킹 방법에 대한 이해! (0) | 2020.07.17 |




댓글